Main page >>
Moulds and their characteristics >>
How moulds are classified >>
Where moulds are found >>
How moulds can be isolated >>
How moulds are grown >>
Contamination >>
Moulds under the microscope >>
How moulds are identified >>
Index of descriptions >>
Bibliography
WHERE MOULDS ARE FOUND
Almost any natural material, no matter how small, will support an indigenous population of moulds. Moulds are part of the economy of nature, quickly occupying dead or nearly dead materials and returning them to the basic building components of new organisms. As such, they are essential to what biologists call nutrient cycling, the process whereby nutrients never leave the realm of living things, but simply get used again and again; earth to earth, ashes to ashes, dust to dust.
While acknowledging that moulds are everywhere, it is useful to classify their habitats into a number of categories based on nutritional characteristics. Moulds have specialized nutritional requirements and usually do not range very far from their usual habitats: fungi that naturally decay seaweed on the beach will not be expected to occur on mouldy bread in the kitchen. The competition among moulds having similar nutritional requirements is intense and leaves no room for a mould adapted to another habitat.
Aside from nutritional and competitive barriers, other factors encourage habitat restriction among moulds. Most influential is spore dispersal. Although most moulds seem to produce astronomical numbers of spores, they are, in fact, practising a strict economy. In their highly competitive life there can be little waste; every minute bit of energy must be put to good use or a more efficient organism will prevail. Thus moulds will produce only enough spores to ensure reproduction of their species from year to year. If a mould were to put a disproportionate amount of energy into spore production, it would have to be at the expense of some other activity, such as rapid growth. Economy in spore production is best ensured by mechanisms of spore dispersal conferring a high probability of encountering suitable places to germinate and grow. Many different mechanisms exist and account for the great variety of spores and spore-producing structures found in moulds.
In many instances, moulds living together in particular habitats have similar methods of spore dispersal, even when the moulds themselves are not closely related, illustrating the idea that many structures in organisms are similar because of common ecological pressures rather than common ancestry. Thus we come again to a basis for subdividing habitats, and, as we shall see later, also a basis for applying certain techniques for the isolation and cultivation of moulds.
Living plants

Most plant diseases are caused by fungi. The largest group of plant parasites are the rust fungi, a highly specialized group of organisms that grow extremely slowly in the laboratory and are outside the usual definition of moulds. Powdery mildews, like the colonies of Uncinula circinatum parasitzing a maple leaf at right, have never been cultivated away from their plant hosts. Some plant disease fungi are less difficult to grow but are so specialized as parasites that they seldom occur in laboratories other than those of the plant pathologist. Many of these fungi have rather strict nutritional requirements to induce sporulation and are usually seen in cultivation only as sterile hyphae.
The plant disease fungi most commonly encountered as moulds are those having non-parasitic stages in their life-cycles. These fungi usually grow and sporulate well in the laboratory. Examples are species of Phoma, Fusarium, Bipolaris, Graphium, Pestalotiopsis, and Monilia.
Most parasitic fungi do not produce great numbers of spores and it is likely that they are transferred to uninfected plants by fairly specific means. Those having wet spore stages, such as species of Phoma, Fusarium, Pestalotiopsis, and Ustilago (a smut fungus), are probably transmitted by insects or other arthopods, such as mites. Other fungi, such as Cladosporium and Monilia, produce large numbers of dry spores, and appear to be wind-dispersed.
Also conspicuous are fungi attacking plants weakened by senescence or disease. Such invaders are hardly parasites in the strict sense. Many of these fungi are now considered to be "endophytes", organisms that enter plant tissues while they are still healthy and remain there in a dormant state until the tissue is weakened or dies. Some ecologists now view these endophytes not as parasites but as the first arrivals in a long sucession of decay organisms. Many species of Alternaria, Botrytis, Cladosporium, Fusarium, and Ulocladium may belong here.
Dead plant material
Herbaceous plants
Moulds occurring on dead herbaceous plants are often the same species attacking dying ones. Thus we can again list Alternaria, Cladosporium, and others, as well as a few new ones such as Epicoccum and Candida. In temperate climates, where many plants die in the fall, there is a tremendous flush of mould activity in autumn. Because the dead plant material is so abundant, finely tuned dispersal mechanisms are not necessary and the moulds simply release their spores in the wind. The likelihood that a spore of Cladosporium, for example, will hit a dead plant part in the autumn is very great.
Nutritional specialization dictates that different plants will support different fungi, and we find, for example, dead grasses yielding a different flora of moulds than dead milkweeds or mustard, at least to a partial extent.
Wood
Dead wood is a good source of moulds and provides two distinct habitats, depending on whether it is covered with bark or not. Moulds occurring under the bark of fallen and standing trees cannot disperse their spores into the air and most often utilize insects and mites for their transmission. Although, as we might expect, many have wet spores ideal for sticking to the bodies of their dispersers, some have dry spores that appear to become trapped among the animals' body hairs. I have seen hundreds of the curved, dry ascospores of Fragosphaeria purpurea trapped among the bristles on the backs of mites under bark of maple trees in Ontario. Often the moulds under the bark of dead standing trees are different from those on fallen trees.
The surface of wood not covered with bark often has a distinct community of moulds. Although air dispersal of spores is possible in this situation and apparently often occurs, wet-spored moulds are still abundant. Again standing and fallen trees support different fungi. Freshly cut wood is a good source of moulds causing a phenomenon called blue stain. These moulds, common in lumber-yards as bluish to black discoloration on the wood, produce tall sporulating structures bearing wet drops of insect-transmitted spores. Closely related species occupy the tunnels and galleries formed by beetles under the bark of living and recently dead trees.
Leaves
Dead or dying leaves of trees support moulds similar to those from herbaceous plants, with some notable exceptions. However, tree leaves appear to support fewer Alternaria and Cladosporium colonies, thus often allowing a better chance to observe the more slowly growing or rarer fungi. For example, in Ontario fallen leaves of ash support a pin-sized mushroom called Marasmius minutus, that never occurs on herbaceous plants.
Leaves submerged in water support a number of moulds with unusual spore forms. The spores may be needle-like, coiled like a watch-spring, or coiled into barrel-shaped structures that bob to the surface of the water and float away. Others have three or four narrow arms and look something like jacks with two or three of the arms missing. These moulds may be examined by submerging leaves in a bowl of water and stirring them vigorously. After a few minutes the spores will float to the surface, where they can be skimmed off in a drop of water. Best results are obtained with leaves likely to become submerged later in the natural habitat but have not yet become so.
Animals and humans
Diseases
A number of diseases of humans and animals are caused by mould and yeast-like fungi (Figure 7). Many are known only from this habitat and are quite specialized. Notable are the moulds called dermatophytes, the cause of a number of skin diseases such as ringworm and athletes' foot. The fungus grows on the outermost layer of skin, causing reddening of the surrounding tissues (zoophilic types) and sometimes scaliness (anthropophilic types). Dermatophytes do not normally attack deeper tissues; the symptoms are usually due to an allergic reaction. Because of their essentially non-parasitic nature dermatophytes are usually easy to grow in the laboratory. Many closely related species occur on substances similar to human skin, such as leather, feathers, hair, and horn, and are unable to grow on living animals or man. Probably the most commonly isolated dermatophytes are species of Microsporum (Figure 7A), Trichophyton, and Epidermophyton.
Several fungi can occur on living tissues and cause serious disease and even death. Many such organisms are known only from individuals who have low disease resistance, due to prior infection, AIDS, old age, or other factors. Most commonly, infections by these fungi occur when the normal bacterial populations of the body are eliminated by the use of antibiotics. Without competition from bacteria these fungi occupy the tissues and grow rapidly, often causing considerable damage.
Other fungi attack healthy living tissues without the aid of antibiotics, causing more localized but very serious infections. Histoplasmosis, a disease with symptoms similar to those of tuberculosis, is caused by Histoplasma capsulatum (Figure 7C), a mould associated with bird nests in nature. It is frequently reported in perfectly healthy people who have been exposed to the dust from nesting materials, such as while tearing down old barns.
The study of medically important fungi is beyond the scope of this book and will not be discussed further. There are several good books dealing with medical fungi from a mycological (rather than medical) point of view. Especially useful are Hoog and Guarro, 1995; McGinnis (1980) and St-Germain and Summerbell, 1996.
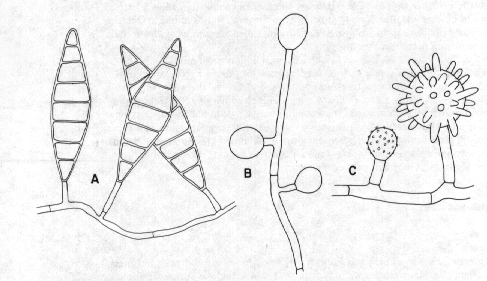
Figure 7. Some fungi of medical importance. A: Microsporum, causative agent of ringworm and other skin diseases. B: Blastomyces dermatidis, causative agent of North American blastomycosis. C: Histoplasma capsulatum, causative agent of histoplasmosis.

One of the most devastating animal diseases to have appeared in recent years is the "White Nose Syndrome" in hibernating bats. This disease, caused by Geomyces destructans is now known from a large part of eastern North America where it has drastically reduced the numbers of bats. Some scientists have suggested that species such as the little brown bat may become almost extirpated in much of its geographical range. The fungus appears on the hibernating bats during the winter, causing them to awaken several times. It is thought that when they wake up they burn of valuable fat reserves necessary to carry them through until the return of flying insects in the spring. Infected bats attempt to leave their caves to feed in late winter and freeze or starve to death. The photo at right shows two infected bats photographed in an old mine in New Brunswick, Canada. The fungus can be seen at white circles on each side of the animals' noses.
Predation
Some moulds actually trap small (usually microscopic) animals. Best known are those that trap eelworms or nematodes (see Barron [1977] for an excellent account of these fungi). The traps produced by these fungi are composed of loops, networks of filaments, or knobs and branches. Some are reminiscent of the well known Venus' fly-trap, and are sprung upon contact with an eelworm. When the worm crawls into a ring of one of these fungi, the ring rapidly constricts, holding the eelworm tightly. At first the worm struggles violently, but it soon becomes still and is invaded by the filaments of the mould. In other species, the loops and branches are sticky and hold the eelworm fast as soon as it touches them.
A second group of eelworm catchers produces spores that are swallowed by the worm, germinating later in the gullet. At least one species has an elaborate injection mechanism that, when sprung, forces a small portion of the spore right into the tissues of the side of the eelworm.
Soil
Soil is one of the most commonly studied mould habitats. It is source of plant disease fungi, some human disease fungi, predacious fungi, and a host of forms that bring about the transformation of dead plant and animal material into soil.
Soil is not a uniform substance; in fact, it is so complex that a study of soil is a science in itself. Scientists recognize several layers in the soil, starting at the surface and working down. These are:
L layer: the layer at the soil surface composed of undecayed plant litter. The origin of the litter is clearly identifiable.
F layer: the layer with identifiable but partially shredded or decayed plant material.
H layer: the layer containing much material of plant origin, but with individual structures no longer recognizable.
A layer: the layer with plant or organic matter present but unrecognizable as such or well mixed with loam (a mixture of sand, silt, and clay).
B layer: a mineral layer containing little organic matter.
C layer: the parent rock from which the mineral layer is derived.
Naturally these layers are not always distinct, and grade more or less evenly on into the other. Some soils, especially agricultural ones, lack stratification altogether. Soils differ from one another in the kind of plant material falling into them; the relative proportions of sand, silt, and clay in the mineral components; the moisture content; the relative acidity or alkalinity (pH); and in many other characteristics.
Fungi, because they are highly specialized users of organic substances, vary from one soil to another and with depth in the same soil. They are entirely dependent upon the kind of material falling into the soil: a fungus specializing in oak leaves will probably not colonize a pine needle. This factor alone will account for many differences between the mould populations of two soils. In addition fungi may be sensitive to moisture levels, pH, competition from other organisms, and many other influences.
As a leaf falls into the soil it is shredded by arthropods and digested by microorganisms. Gradually it becomes buried under newer generations of leaves and thus moves down into the lower layers of soil. It undergoes continuous decay by moulds, bacteria and other organisms until it finally disappears (somewhere in the B layer). Fungi that first invade the leaf, often while it is still on the tree, will grow on it until the particular nutrient they need is exhausted and then die back, to be replaced by new fungi extracting what they need from the leaf. In this way, a leaf moving down through the layers of soil will have a series of moulds on it, each replacing a previous population. This transition of populations as the habitat ages and is modified is called succession and is the object of much study and discussion among ecologists.
Although we tend to think of soil fungi as decomposers of litter, they play a number of other roles as well. Many are associated with animals and animal products and may trap eelworms and other soil animals, or decay the dead bodies of insects and earthworms. Some invade cast-off feathers and hair, and at least one group specializes in old hooves and horns.
Through their roots, living plants offer an attractive habitat for soil-inhabiting fungi. Some live around the root and decay away the dead root layers or substances exuded by the root. Others invade the living root tissues and either cause plant disease or live in harmony with them. The latter situation involves a form of symbiosis called mycorrhizae, where the root and the fungi within each benefit the other, In fact, most mycorrhizal fungi and plants cannot live apart. Only a few species of plants are consistently free of this association.
Air
Air, of course, is not a habitat for moulds, though many disperse their spores by air currents and are encountered in routine work.
Fungi have a variety of mechanisms for getting their spores into the air. Simplest is that of exposing dry spore masses to air currents. Many moulds use this method, especially those that colonize exposed leaves and stems. Species of Alternaria, Cladosporium, and some of the basidiomycetes called smuts are conspicuous for this kind of dispersal. These species usually produce large numbers of spores, a necessary expenditure if at least a few randomly dispersed spores are to land on a suitable place to grow.
Ascomycetes and basidiomycetes are usually capable of shooting their spores away into the air. Asci are like popguns: the water pressure inside the ascus builds up higher and higher until the spores are fired out the end with considerable velocity. Ascospores can, in fact, be shot a distance of several centimetres from the end of the ascus, a prodigious feat considering the microscopic size of these mechanisms. The equivalent at our own scale would be to fire a 30-cm projectile a distance of 6 km, a feat necessitating a 105-mm howitzer! With such a mechanism, ascomycetes need not be exposed directly to the wind but may grow in places allowing entry to the air of their discharged spores. Thus ascomycetes are often found growing closer to the ground than other air-dispersed organisms.
Basidia discharge their spores from the tips of small projections called sterigmata. Such spores, sometimes called ballistospores, are not shot very far, only far enough to clear the surface of the basidium and the fruiting structure. The fruiting structures of basidiomycetes may be microscopic in size and bear only a few basidia or they may be very large, as in mushrooms and bracket fungi. These last fungi often continue to produce spores throughout the growing season and can produce a staggering number. One of the most prolific of these, the "artist's fungus", Ganoderma applanatum, may disperse its airborne spores many kilometres from the woods where it grows. Related to the basidiomycetes are the mirror yeasts, a group of yeasts that produce ballistospores on solitary sterigmata (also see discussion of ballistospores in Tremella encephala. Most basidiomycetes are not normally encountered as moulds, and are really outside our interest here, but a few, such as Sistotrema brinkmanii may be encountered in the laboratory.
Some spores become airborne by a mechanism known as droplet adhesion, a process dependent upon the presence of tiny water droplets in the air. When one of these droplets, as on a foggy day, encounters a mould spore attached to a leaf, for example, the spore adheres to the droplet by surface tension and is carried off on it. It is then either deposited when the drop comes to earth, or becomes truly airborne when the drop dries out.
There are a few other mechanisms whereby spores become airborne, but these are rather more specific to one or a few species. Most are forcible discharge mechanisms and may involve various kinds of spring, water jet, or bellows devices. A discussion of these and other common mechanisms is found in the excellent book on dispersal in fungi by C.T. Ingold (1953).
Dung
Although people expect dung to be a rather disgusting material to study, many become so intrigued with the moulds and other organisms found there that they soon overcome their initial objections.
Many fungi found on dung are highly specialized for growth there and never occur anywhere else. They do not produce large numbers of spores and thus require a high probability of success in getting their spores from one dung pile to another.
The most commonly cited life-cycle in dung-inhabiting or coprophilous fungi is that shown by many ascomycetes. The spores of these ascomycetes are very heavy, sticky, and dark, and when they are shot from the ascus their weight allow them to travel a relatively great distance. Because of this weight, however, they do not usually remain in the air long, but follow a parabolic trajectory and land near the dung pile. Because they are sticky, often with rather elaborate slimy coverings or tails, they adhere to whatever they fall on, and because of their dark colours they are not greatly affected by the ultraviolet radiation in the sunlight to which they are exposed for an often considerable length of time. Eventually they may be eaten, if they have fallen on vegetation, by a grazing animal, ultimately to be packaged up in a new pile of dung. Many of these spores have a dormancy mechanism that prevents them from germinating on the vegetation but that can be broken when subjected to the processes of the animal's digestive tract. Most coprophilous ascomycetes are light-sensitive and aim their spore shots away from the dark-coloured dung, a first step in their journey to a new dropping.
Other coprophilous moulds present their spores to the environment in wet drops at the ends of stalks. When one of the many insects or mites attracted to dung brushes against these stalked spore drops they carry with them a few of the sticky spores. Later, perhaps on a new dung pile, the spores rub off and germinate to produce a new colony. Most insect-transmitted fungi lack dormancy mechanisms in their spores, and are unlikely to be away from dung for more than a few hours.
The variety of coprophilous fungi is large, and many occur in the laboratory as moulds. Included are several kinds of ascomycetes, some small mushrooms, slime moulds, zygomycetes, and many anamorphs. It is a particularly rewarding group for study by the beginning mycologist.
One warning must be issued. Dung may contain the eggs, larvae or adults of parasitic organisms. While most of these are of danger only to the species of animal producing the dung, some are able to parasitize humans. Because of this dung should never be handled with bare hands.
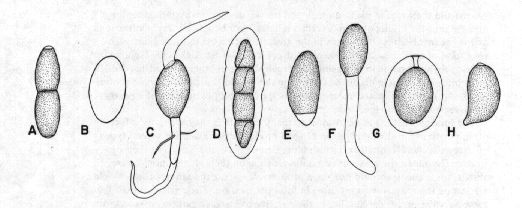
Figure 8. Spores of some dung-inhabiting fungi. A: Trichodelitschia. B: Thelebolus. C: Podospora. D: Preussia. E: Triangularia. F: Cercophora. G: Sordaria. H: Coprinus.
The human environment
Food
Aside from their role in plant, animal, and human disease, many moulds enter directly into human affairs, in either a harmful or beneficial way. On the plus side is the involvement of moulds in the greatest contribution to medicine of all time, that of antibiotics. The discovery of penicillin by Sir Alexander Fleming in 1928 probably resulted in the saving of more lives than all other medical discoveries combined. Penicillin, a product of the common mould Penicillium chrysogenum, is still one of the safest and most widely used of antibiotics, in spite of a more than fifty-year search for others.

The foods we eat are as nutritional to many moulds as they are to us, a fact often put to use in the preparation of food products. For example, several types of cheese, such as Roquefort, Danish blue, Camembert, and Brie, owe their distinctive flavour to the presence of mould growing on them. If the mould were absent, these cheese would not ripen properly. Yeasts, although not really moulds, are among the most important fungi in food preparation. Their value, at least for some Saccharomyces species, lies in their ability to produce carbon dioxide and grain alcohol. In wine-making, where the production of alcohol is valued, yeast is added to the grape juice to bring this about. In bread-making, the important product is carbon dioxide, which is necessary in the rising process, and, again, yeast is added to the dough. In the production of beer, both alcohol and carbon dioxide, to produce carbonation, may be necessary, although today the carbon dioxide may be added later artificially.
In the Far East, a number of moulds are used in food preparation that remain unexploited in most of the world. Among these are species of Aspergillus, Monascus, and Rhizopus, used to process various rice, bean and soybean products.
To most of us, the negative aspects of moulds in foods are more noticeable than the positive. Few of us have failed to notice the pink, black, and green moulds growing on bread or the green or blue-green Penicillium rots of citrus fruits. Moulds are one of the reasons food manufacturers include preservatives in their products. The picture at right shows a species of Penicillium, growing over the surface of some tomato sauce left too long in the refrigerator. The yellowish drops of liquid, called exudate, resting on the surface of the mould, and its overall blue-green colour suggest it is P. chrysogenum. Microscopic examination revealed it had smooth conidiophores and conidia, also indicating it to be P. chrysogenum.
One of the most destructive activities of moulds in foodstuffs occurs in stored seeds and grains. Certain species of Aspergillus and Penicillium are able to grow under particularly dry conditions and attack stored grains. To guard against this, the grain must be dried to very low moisture levels. Keeping grains dry in the humid tropics is particularly difficult if not, at times, nearly impossible. Not only do such fungi damage the grains or render them unpalatable, they may also excrete toxins that can cause illness or even death. Most famous of these are the aflatoxins - produced by Aspergillus flavus and other moulds - which are not only immediately toxic but are also known to be carcinogenic. Aspergillus flavus often grows on peanuts and was first discovered in peanut feed given to turkeys in Great Britain.
Some of the same species of Aspergillus that attack stored grain commonly appear on the surface of jams, jellies, and syrups. These very sugary substances tend to prevent water from entering the cells of most fungi and thus create what amounts to a microbiological desert. Aspergillus species, with their ability to grow under dry conditions, are ideally suited to such habitats. Some products, such as prunes and dried apricots, are so dry and so sugary that even Aspergillus species are unable to cope with them. They still can be decayed by certain specialized moulds, however. Probably the most remarkable to these is Xeromyces bisporus, which cannot even grow at the sugar concentration used to grow Aspergillus species. Laboratory cultures of this species are rather difficult to establish and require very special conditions.
Cellulose products
Cellulose is probably the most abundant material of biological origin on the earth and is a prime source of energy for many fungi. We have met some cellulose-decomposing fungi already in our discussion of moulds inhabiting dead and dying plant parts, notably Alternaria, Cladosporium and Epicoccum. While these fungi are common enough, there are several others that seem to become especially common on man-made cellulose products, such as paper, cotton, cardboard, and wood products. Many mould growths on paper and cotton are commonly called mildews, but this term really has little meaning. These fungi are able to dissolve the cellulose fibres in cotton and paper and thus cause the materials to disintegrate. The process is especially rapid under moist conditions, as occur in damp basements or in the tropics. Many species of moulds are involved in cellulose decomposition, but none are more widespread than species of the ascomycete genus Chaetomium. During the Second World War, countries fighting in the South Pacific and Southeast Asian areas lost a great deal of equipment to these species. The resulting increased interest in Chaetomium is reflected most notably in Ames's (1963) monograph on the Chaetomiaceae, published by the United States Army.
Sawdust contains many cellulose-utilizing moulds that may cause unwanted decomposition. Particularly undesirable are members of a group of moulds able to grow at high temperatures (up to 60░ C). The growth of these moulds may actually cause the temperature of the sawdust to rise. Combined with bacteria having an even higher temperature tolerance and natural chemical activities these moulds may eventually lead to spontaneous combustion, a common cause of fire in lumber-yards.
Other products
Many other manufactured products are susceptible to mould attack. Painted walls, particularly in humid places such as showers, can become overgrown by certain fungi, notably species of Phoma and Exophiala. Wallpapers also serve as a source of nutrition for some moulds. Commonly cited are the Scopulariopsis species which have been reported to grow on wallpapers containing arsenic pigments and to release very poisonous gases. With modern wallpaper pigments, which contain no arsenic, this cannot happen.
Leather is mainly protein and serves as a convenient source of food for moulds. Some of the moulds that occur here are related to the dermatophytes that attack the outer layer of human skin; others belong to quite different groups.
Some substances that seem to be entirely unlikely to support fungal growth may be found to be well colonized by these organisms. Exophiala species, for example, are found in syrupy solutions of polyvinyl alcohol. Cladosporium (Amorphotheca) resiniae often occurs on the surface of aircraft fuels in their tanks and can damage jet engines. Penicillium ochrochloron can be found in electroplating solutions that are extremely acid and contain very high levels of toxic copper salts. I was once involved in a search for the organism responsible for decaying inflatable life-rafts periodically soaked in seawater. The culprit eventually turned out to be a species of Aspergillus.
Moulds in the indoor environment
Increasing attention is becoming focused on fungi in indoor environments. Although most of the moulds occurring in human environments occur indoors, the interior of buildings are themselves a special habitat. It has long been known that indoor fungi cause allergies in sensitive persons, but only relatively recently have indoor moulds been linked to other health problems. While allergy sufferers generally react to substances in fungal spores, other health problems may be caused by voltile substances released by moulds into the air.
Virtually all buildings contain moulds, but some are mouldier than others. Indoor moulds can be remarkably tolerant of dry conditions but none can live without some moisture. Excessively mouldy buildings generally have a source of moisture leading to unusually heavy mould growth. The source of the moisture may be a leaky basement, a dripping pipe, a roof in need of repair or some other fairly obvious cause. In most cases the mould can be seen growing on walls or other materials in contact with the moisture. Sometimes the moisture can occur inside walls and not be apparent. A common but not obvious cause of moisture in cold climates is condensation inside north-facing walls. Severely mouldy buildings may have a musty smell, but not necessarily. Sometimes the only sign of a problem is persistant poor health of the occupants, such as headaches, nausea, respiratory symptoms, etc. It is now known that mouldy buildings can present a serious health risk to occupants. In fact, some infant deaths have been convincingly linked to indoor moulds. Any building with an apparent mould problem should be thoroughly investigated by qualified people.
Not all indoor moulds present a risk to human health, but an abundance of any mould is likely to be accompanied by others, including toxic ones. Species of Stachybotrys are particularly toxic.
Obvious occurrences of Stachybotrys may be sufficient cause for a major "decontamination" by qualified technicians wearing special protective clothing.
 Stachybotrys species produce black colonies on dry wall, ceiling tiles and other materials containing cellulose. The easiest way to confirm the presence of Stachybotrys is to press a piece of cellulose tape against the mouldy spot and then examine it sticky side up on a slide under a microscope for the characteristic spore-bearing structures. The image at right illustrates conidiophores and conidia of a Stachybotrys species collected on tape from a basement in southern Quebec. Not all moulds are identifiable using this technique, but it works well for many, including Stachybotrys.
Stachybotrys species produce black colonies on dry wall, ceiling tiles and other materials containing cellulose. The easiest way to confirm the presence of Stachybotrys is to press a piece of cellulose tape against the mouldy spot and then examine it sticky side up on a slide under a microscope for the characteristic spore-bearing structures. The image at right illustrates conidiophores and conidia of a Stachybotrys species collected on tape from a basement in southern Quebec. Not all moulds are identifiable using this technique, but it works well for many, including Stachybotrys.