Home >> Where fungi grow >> Mutualists >> Mutualisms with plants >> Mycorrhizae
MYCORRHIZAE
Mycorrhizae (singular: mycorrhiza) are mutualisms formed between fungi and plant roots. The importance of mycorrhizae cannot be overstated; it has been suggested that as many as 95% of all the world's plant species form mycorrhizal relationships with fungi and that in the majority of cases the plant would not survive without them. Mycorrhizae have existed for a very long time and can be demonstrated in the fossilized roots of some of the earliest land plants. They can be found in plants growing in habitats ranging from humid to dry tropics all the way to the far north and south. Some scientists have suggested that plants were only able to move on to land when they had developed mycorrhizal relationships with fungi.
Mycorrhizae are considered to be a mutualistic relationship because both organisms benefit. The fungus receives the products of photosynthesis from the plant and is thus freed from the necesity of finding its own sources of energy. At the same time the fungus grows out into the soil and retrieves nutrients, especially phosphorus and nitrogen, and passes these back to the plant. Numerous experiments have shown that plants without mycorrhizae cannot cope as well with low mineral levels as those that have mycorrhizae.
It is generally believed that mycorrhizae have been re-invented many times over the history of land plants. Many different groups of fungi are involved and the form of the actual fungus-root interface, the mycorrhiza itself, varies greatly. Scientists recognize several distinct types of mycorrhizae and can relate these to particular groups of plants and fungi. The most thoroughly studied of these types are arbuscular mycorrhizae, ectomycorrhizae, ericoid mycorrhizae, arbutoid mycorrhizae and orchid mycorrhizae
Arbuscular mycorrhizae
Arbuscular mycorrhizae (often called AM) are the most common and widespread of all mycorrhizae and are found in as many as 85%-90% of the world's plant species.
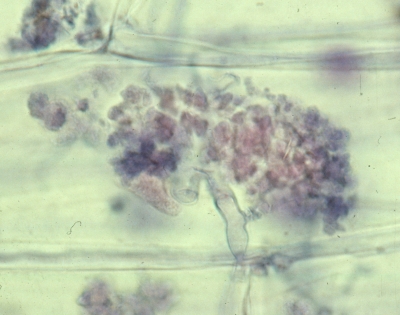
In this association the fungus occurs inside the cells of the plant root as a highly branched shrubby structure called an arbuscule. The picture at right shows an arbuscule of one of these fungi inside a root cell of a small onion plant. To view this fungus is was necessary to clear all of the contents out of the root cells with hot potassium hydroxide followed by a blue dye called Trypan Blue. The cellulose of the root tissue did not stain very much while the wall material of the fungus, possibly chitosan, stained purple. Although it appears that the arbuscule fills the root cell it actually occurs between the root cell wall and the cell membrane within. The cell membrane fits over the arbuscule like a rubber glove over your hand. Thus the fungus never comes into direct contact with the root cell nucleus, mitochondria or other cell structures. The exchange of nutrients between the two partners, minerals from fungus to plant and sugars from plant to fungus, takes place at the cell membrane-arbuscule interface.

Fungi forming arbuscular mycorrhizae may produce some other structures in addition to arbuscules. Roots harbouring arbuscules may also contain balloon-like structures called vesicles as illustrated above in the root cells of the same onion shown previously. Vesicles are very commonly produced by these fungi, so often, in fact, that they have been referred to as "Vesicular-arbuscular Mycorrhizae (VAM)", particularly in the older literature. However, that name is unnecessary because these fungi nearly always produce arbuscules but may often lack vesicles.
The fungi forming arbuscular mycorrhizae not only occur in roots but also extend out into the surrounding soil where they form a dense network. It is this so-celled extra-radicular phase that is responsible for extracting nutrients from the soil. The hyphal network is relatively long-lived and is able to colonize new roots as they enter its domain. One of the criticisms of soil tilling, a common agricultural practice, is that it breaks up these mycorrhizal networks and prevents germinating seeds and young plants form tapping into existing systems of nutrient uptake.
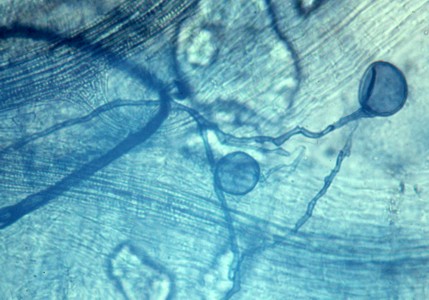
Reproduction of the fungi forming arbuscular mycorrhizae is by thick-walled spores produced on the extra-radicular hyphae. These are thick-walled and often remain in the soil for long periods. Many are quite large and can be recovered by sieving the soil. Although these spores will germinate and produce hyphae in the laboratory no one has yet succeeded in growing the fungus independently of a root. The picture at right shows spores of a species of Glomus produced on a branched extra-radicular hypha.
The fungi able to form arbuscular mycorrhizae are relatively few in number and all are members of their own phylum, the Glomeromycota. Although the root system of a plant might support several species of these fungi the diversity is never great. In fact some species appear to be common throughout the world, in spite of immense climatic and floristic differences.
Ectomycorrhizae
Ectomycorrhizae, sometimes abbreviated as EM, are not as widespread in nature as arbuscular mycorrhizae, occurring on the roots of about 5% of the world's plants. The most diagnostic feature of ectomycorrhizae is that the fungus never penetrates the cell walls of the plant and thus the exchange of nutrients must take place not only through the cell membrane but through both plant and fungal cell walls as well.
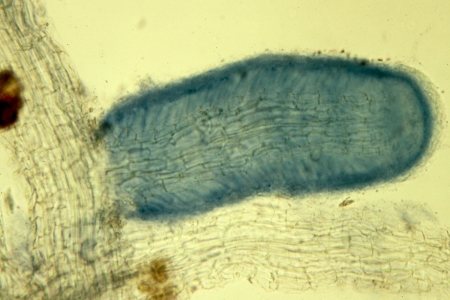
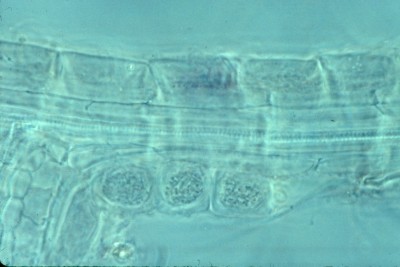
A young root of paper birch is shown in the illustration at right. It was stained with Trypan Blue as outlined above and shows an older section uncolonized by fungi and thus unstained giving rise to a new branch that has become heavily colonized. The outside of the root is ensheathed in a thick covering of hyphae called a mantle, visible in the picture as a dark blue border around the root. The mantle is outside the root and can often be removed as though it were a shell. It completely isolates the root from its environment so that all substances coming into that root must come from the fungus. Inside the mantle is a single layer of cells forming the epidermis, the outermost tissue of the root. In paper birch and several other species the epidermal cells become elongated and slanted to form a herringbone pattern following fungal colonization. Below the epidermis is a layer of cells forming the cortex of the root. The cortex varies in thickness according to the species of plant, ranging from one to several cells deep. The colonizing fungus grows between the cells of the epidermis, and often the cortex as well, and surrounds the individual cells, finally isolating them from one another. This network of enveloping hyphae surrounding epidermal and often cortical cells is called the Hartig net in honour of the nineteenth-century German biologist Robert Hartig who first observed it. The Hartig net is the interface between the fungus and the plant; it is the site of all nutrient and water exchanges between the two associates. Although the Hartig net surrounds the plant cells they do remain in contact with one another through microscopic strands called plasmodesmata. The centre of the root is taken up by the stele, the conducting system of the root. Water and nutrients passed by the fungus to the root are transported throughout the plant via the stele, itself composed to several parts.
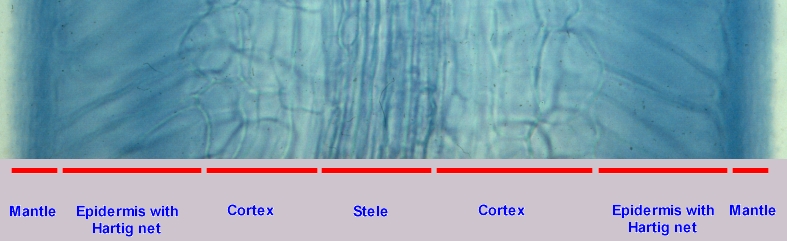
The picture at right illustrates these relationships more clearly. It presents a narrow strip across a mycorrhizal root of Populus tremuloides, the trembling aspen. Comparing this picture with the one above of paper birch you can see that the root tip would be at the bottom of the page. At this higher magnification the compactly arranged hyphae of the mantle can be seen as well as the hyphae of the Hartig net surrounding the slanted epidermal cells. The cortex can be seen to be made up of more or less round cells while the stele is clearly formed of elongated cells. In the aspen and birch roots the Hartig net extends only into the epidermis, but in other species, white spruce for example, the cortex may also become involved. The location of the fungus within the root is under the control of the plant, which uses toxic phenolic compounds to limit growth of the hyphae. This can be observed in laboratory experiments where the same species of fungus is introduced into the roots of two species of plants; in one it may extend down into the cortex while in the other it may be restricted to the epidermis.
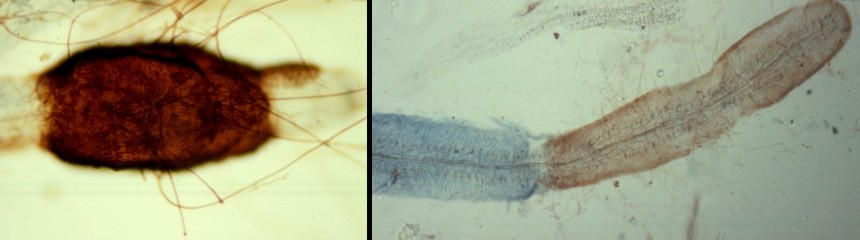
Unlike the fungi forming arbuscular mycorrhizae, those forming ectomycorrhizae are exceedingly diverse. Several thousand species of these fungi are known and many are undoubtedly still waiting to be discovered. With such a diversity of fungi on hand, the root systems of large ectomycorrhizal plants may support dozens of fungal species. Moreover, at least once, and possibly more times in a year the root grows out from its mycorrhizal sheath and must be recolonized. The picture at far left shows an aspen root growing out through the end of a mycorrhizal sheath, probably produced by the fungus Cenococcum geophilum. Although a little out of focus you can see that the emerging root tip is uncoloured and unstained, indicating that it was yet to be colonized. The picture in the right panel shows an aspen root that has had three such growth spurts: the first of these resulting in colonization by a lightly staining but orginally colourless fungus, followed by two additional colonizations by a brown fungus. It appears that the root was orginally colonized by the blue-staining species but that this was replaced by another species following a growth spurt. The third growth spurt simply resulted in recolonization by the brown species. These events suggest that as a root grows it may continue to associate with one species of fungus but it also has the opportunity to change partners. Numerous studies have shown that ectomycorrhizal fungi differ greatly from one another in their physiology and it is tempting to think that as a root extends out through the soil it is able to form an association with a fungus apprpriate to the particular conditions it encounters.
The ability to form ectomycorrhizae is found in many families of fungi, but most commonly among members of the class Agaricomycotina of the Basidiomycota, especially those producing mushrooms and boletes. Most of the larger mushrooms you see in the forest have arisen from the networks of extra-radicular hyphae permiating the soil beneath your feet. The abundance of these mushrooms, their sheer weight and volume, attests to the magnitude of their activities. The energy and chemicals needed to build these mushrooms comes in great part from the trees, suggesting that the advantages a plant gains from mycorrhizae come at a cost.
Most plants exclusively form arbuscular mycorrhizae but there are compelling reasons to focus attention on those having ectomycorrhizae as well. Although a smaller number of species are involved, ectomycorrhizae dominate in the pine, oak, birch, willow, walnut and several other families. In the tropics these include the dipterocarps and large woody legumes. In New Brunswick our extensive forests of spruce, fir, white pine, birch and poplar support immense continuous networks of ectomycorrhizal fungi. Without these fungi our forests as we know them would not exist. Thus the ecological and economic importance of ectomycorrhizae cannot be overestimated.

Many biologists have noted the major differences between tropical and temperate forests and have attempted to relate these to dominance by certain mycorrhizal types. The pictures above illustrate two such forests; at left a tropical rain forest in northern Costa Rica and at right a forest near Schefferville, Quebec. The Costa Rican forest is dense and made up of a great variety of tree species. You might walk some distance through this forest before encountering two individual trees of the same species. Biodiversity here, including the trees, seems to be high. On the other hand the Quebec forest appears to have only one kind of tree. Closer examination would reveal some four or five species but hardly more. If you started walking away from the base of a spruce tree it wouldn't be long before you encountered another. Biodiversity here seems to be very low. Curiously, fungal biodiversity in these forests takes another form, at least when it comes to mushrooms. Few trees in the Costa Rican forest are able to form ectomycorrhizae while in the Quebec forest all the trees form ectomycorrhizae. As a result of this the Quebec forest will have a great variety of large mushrooms while the tropical one will support a lessor variety of mostly small mushrooms.
Other kinds of mycorrhizae
Although arbuscular and ectomycorrhizae account for most instances of mycorrhizae there are some other more specialized types. These are all restricted to a single plant family, or a closely related group of families, and are not as commonly encountered as the two main types. However, in New Brunswick we are fortunate to have these other types of mycorrhizae and can observe them during the active growing season.
The most widespread of the "minor" groups of mycorrhize are ericoid mycorrhizae, so named because they are restricted to the Ericaceae, the heath family, and some closely related families. The Ericaceae are familiar to New Brunswickers in the form of blueberries, cranberries, sheep laurel, rhododendron, etc. Crowberries, in the related family Empetraceae, also have ericoid mycorrhizae. The picture at right shows a root of Labrador tea, another member of the Ericaceae common throughout New Brunswick. Compared with the birch and aspen roots those of Labrador tea appear to be lacking some tissues, but in fact everything is there. These roots are exceedingly thin and often called "hair roots". If you attempt to pull one of these plants out of the soil without loosening the roots first with a shovel or trowel you will leave all of the hair roots in the ground. In the picture there appears to be only a cortex and a stele but there is also an inconspicuous epidermis that didn't show up very well in the photograph. The cortex is only one cell thick and these cells are packed with fungal hyphae. In many ericoid mycorrhizae the fungus may actually outweight the root! Such a root is little more than a receptacle for holding a fungus. Although the hyphae appear to fill the cortical cells they are outside the cell membrane, which is intricately wrapped around them. Nutrient exchange between the fungus and plant takes place at this interface and is extremely efficient.
The plants forming ericoid mycorrhizae are highly adapted to this association and, not surprisingly, so are the fungi. These fungi, most commonly the inoperculate discomycete Rhizoscyphus ericae, have highly developed abilities to extract nutrients from nutrient-poor soils. Hence members of the Ericaceae are able to grow in such places as bogs and sandy areas that would be unavailable to most plants.
Arbutoid mycorrhizae also occur in the Ericaceae but only on a few species, mainly those of Arctostaphylos and Arbutus. Arctostaphylos uva-ursi or bearberry is the only representation in New Brunswick and it is not very common, although it is abundant to the north and west of us. Arbutoid mycorrhizae are similar to ericoid mycorrhizae in having the cortical cells filled with hyphae but differ in having a mantle as in ectomycorrhizae. The fungi involved are apparently ones that also commonly form ectomycorrhizae.

Monotropoid mycorrhizae resemble arbutoid mycorrhizae in many ways but differ in the appearance of the roots and in the nature of the mutualism. In the pictures above, all of Indian pipe, Monotropa uniflora, we see at left the above-ground part of the entirely white plant. The picture in the middle shows the base of the stems, including the underground root mass, and the one at right provides an enlarged view of the roots. Indian pipe is common throughout Canada and lives as a parasite. Until fairly recently it was believed that this plant attached itself to the roots of other plants where it would absorb their nutrients. It's lack of chlorophyll suggested that it had no existance independently of its plant host. More recent studies have demonstrated that Indian pipe and some other related plants actually tap into the extra-radicular hyphae of fungi forming mycorrhizae with neighbouring trees and encorporate these into its own dense, concentrated mycorrhizal system. From these mycorrhizae the plant can drain sugars and all the other nutrients it requires. From this we see that Indian pipe is not really a parasite of plants but of mycorrhizal fungi.
Orchids form their own kind of mycorrhizae, often for complete nutrition including sugars. Many orchids are without chlorophyll when young. Others lack it throughout their lives. The use of mycorrhizae to offset this lack of chlorophyll is similar to the situation in Indian pipe. The mycorrhizae themselves are similar to ericoid mycorrhizae, although the roots are larger and the colonization less extensive. The fungi involved in orchid mycorrhizae are interesting in that they are close relatives of fungi causing plant disease. Mycorrhizal fungi in other groups are not closely related to parasitic groups and would present no threat to the plant.