Home >> Diversity and classification >> True fungi >> Dikarya >> Basidiomycota >> Pucciniomycotina >> Pucciniales
PUCCINIALES: THE RUST FUNGI

The Pucciniales is one of the largest orders of fungi, probably containing several thousand species. All of them are obligate plant parasites occurring on ferns, conifers and flowering plants. To fully appreciate these fascinating organisms it is necessary to understand their complex life histories, outlined in the Discussion of the Pucciniomycotina. The picture at right is of the crown rust, Puccinia coronata. It causes an important disease of cultivated grasses in many parts of the world. The left panel of the picture shows a cluster of aecia on a leaf of buckthorn, an intermediate host in the life cycle of P. coronata. These are formed early in the season and produce large numbers of orange aeciospores. The function of the aeciospores is to travel to the surface of the definitive host, germinate and colonize its tissues. The definitive host (the one on which sexual reproduction takes place) is cultivated oats in this example but might also be wheat. The definitive host is often called the telial host because it is the one on which telia and teliospores are produced. In the picture the rust can be seen as orange spots on the surface of the oat leaves. The orange colour is mainly due to the masses of urediniospores, indicating that this is a mid-summer stage of the infection. Teliospores are much darker in colour and would be seen at the season's end.
Puccinia, and the related genus Uromyces are the most abundant of all rusts. They occur on a huge number of plants and can be collected anywhere in the world. In our region they are most conspicuous on grasses, sedges and other monocots, nearly always as uredinia and telia. Their intermediate hosts are likely to be found in spring, producing aecia and aeciospores on a variety of monocots and dicots. They are sometimes difficult to identify to species but you can often find success using a type of book called a "host index" where the fungi known to grow on a particular plant are listed. In Canada we use the Compendium of plant disease and decay fungi in Canada by J.H. Ginns, published by Agriculture Canada. In the United States the standard work is Fungi on plants and plant products in the United States by D.F. Farr, G.F. Bills, G.P. Chamuris and A.Y Rossman, published by APS Press. Of course these books are useful in either country and often complement one another, so most mycologists have both in their libraries.
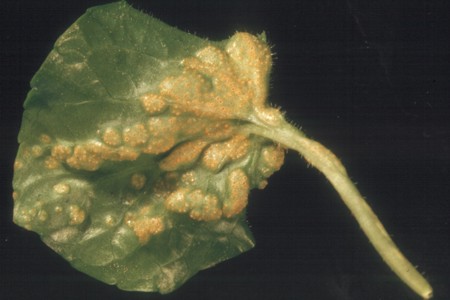

Many rusts are homoxenous (called "autoecious" in rust terminology), having only a single plant host. Thus in the spring you might easily find the rust at left forming aecia and aeciospores on native violets and expect to consult a host index to find out what its definitive host might be, only to be surprised to discover that the violet itself is the definite host. Mycologists have long suspected the ancestors of homoxenous rusts to be heteroxenous (two hosts) and to have dropped one of the original hosts for some reason, a hypothesis supported by recent genetic data. However, you may wonder why an organism that must rely on the successful functioning of five different spore types should manage to survive at all. We are all familiar with the idea that the more moving parts something has the more likely it is to break down. Perhaps the full rust life cycle is vulnerable to such risks. Most of the species of Puccinia on violets have only the one host, but one, P. ellisiana, has Andropogon gerardii, big bluestem, as a definitive host. If the other species of Puccinia on violets descended from ancestors with definitive stages on grasses we might suppose that they gradually became restricted to violets in forest environments where grasses are less abundant and where violet leaves persist from year to year, rust and all. Puccinia podophylli, at right, a species restricted to may-apple, is also a forest-dwelling species that may have followed such a path. This species has also lost uredinia and now depends upon aecia for all its spring and summer reproduction.
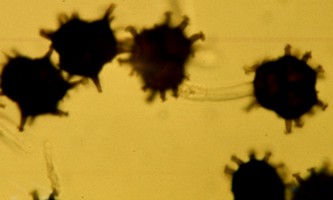
Nyssopsora clavellosa, an autoecious rust on wild sarsaparilla shown at left, has only teliospores (and of course basidiospores). It colonizes the young leaves in the spring with basidiospores produced from the previous year's teleospores. The function of the hooked spines on the teliospores is not known .


Rusts with reduced life cycles can also be found on introduced plants. These may have arrived on our continent with their hosts and survived because they did not require the simultaneous introduction of an alternate host. Two such rusts are seen above. The left picture is a close-up photo of the aecia of Puccinia hysterium, an autoecious rust found on goatsbeard, a plant thought to have been introduced from Russia. The rust in the right picture is Puccinia punctiformis, producing telia and teliospores on Canada thistle, Cirsium arvense. In spite of its name Canada thistle is native to Europe, not Canada.

Some of our most interesting rusts are those growing on trees. These fungi are adapted to the strongly seasonal climate in much of our country and show their greatest variety on fir, spruce, pine, larch and other conifers. With only a very few exceptions these rusts produce spermagonia and aecia on conifers and their uredinia and aecia on smaller understory plants. The large evergreen confers present an ideal platform for the mass release of aeciospores, each ready to colonize the tissues of their definitive hosts. The balsam fir at right has been colonized by Uredinopsis americana and is producing aecia along its needles. The aeciospores will infect newly unfolding fronds of the sensitive fern, Onoclea sensibilis There are four genera of rust fungi producing aecia on balsam fir, Hyalopsora, Melampsorella, Milesina and Uredinopsis, and all have ferns as definitive hosts. Balsam fir needles are the theater for other life and death dramas, often involving insects such as gall midges.

One species, Melampsorella caryophyllacearum, has a definitive host that is not a fern. But it is unusual in other ways as well. Pictured at left is a kind of dense bushy growth on balsam fir called a "witches' broom". This is caused by Melampsorella caryophyllacearum that has colonized the tissues of the tree and chemically induced it to produce the weird proliferation of branches. Many of the branches, such as the one in the right panel of the picture, bear needles on which the aecia are found while many others, produced in previous years, are dead. The definitive hosts of these witches' broom rusts are chickweeds and starworts, members of the genera Cerastium and Stellaria of the family Caryophyllaceae. Witches' brooms are also produced on spruces throughout their North American range but are caused by species of Chrysomyxa rather than one of the rusts known on firs.
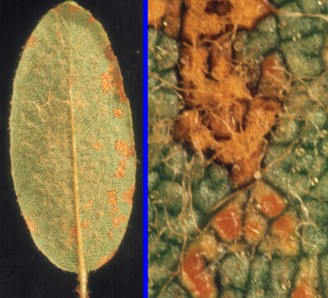

Chrysomyxa species all produce spermagonia and aecia on spruces and have definitive hosts in the blueberry family (Ericaceae). Black spruce can become particularly affected in New Brunswick where small trees in bogs may be orange in colour due to the masses of aecia and aeciospores. Such trees will produce a dusty orange cloud of aeciospores if you shake them. Chrysomyxa ledi, in the pictures at right, is a species commonly found on spruce in our region. Its definitive host, Labrador tea, is very abundant in bogs and is commonly colonized by this rust. The leftmost picture of the pair shows C. ledi on a small black spruce and is recognized by the strongly discoloured needles. An individual plant of labrador tea can be seen in the upper left part of the picture. The panel at far right shows C. ledi on Labrador tea. Most members of the blueberry family can become colonized by rusts and most of these parasites will be species of Chrysomyxa.

Pines are often hosts to species of Cronartium, which produce their aeciospores in crusts and blisters that break through the bark on trunks and larger branches. Some cause the pine to produce conspicuous swollen galls along their branches. These gall- and blister-rusts often kill the branches they are growing on and if severe can kill whole trees. The most infamous of these is the white pine blister rust, caused by C. ribicola a species introduced to North America from Asia via Europe. Its introduction was facilitated by the ability of this fungus to utilize a variety of five-needle pines as aecial hosts and various species of currants (Ribes spp.) as definitive hosts. Cronartium comptoniae, the species illustrated at right, produces aecia on jack- and lodgepole pines. These can occur on the main stem where they appear as large blister-like aecia. The definitive host is Comptonia peregrina, sweetfern, an understory plant common in sandy areas across Canada.
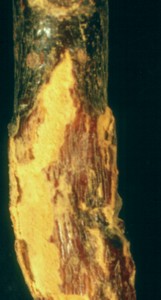
Species of Melampsora differ the tree rusts discussed so far in their frequent choice of trees as definitive hosts. Most have conifers as aecial hosts and either willows or poplars as definitive hosts. The species in the picture at left is producing abundant urediniospores on a branch of a willow. Not all species produce such blisters; most occur on the leaves as small spots. The telia of Melampsora species are produced on the leaves of the willows and poplars as crusts below the epidermis. The teliospores are never released as spores but produce their basidia while embedded in the fallen leaves. Any rust you are likely to encounter on either poplar or willow will be a species of Melampsora and nearly all will have aecial stages on conifers.

The genus Gymnosporangium is composed of species quite different from any of those discussed above. Although gymnosperms are always hosts, none of these are are conifers and none are aecial hosts. Instead, the aecial hosts are all members of the Maloideae subfamily of the rose family, such as serviceberries, mountain ashes, hawthorns, apples and pears. The definitive hosts are junipers. The picture above illustrates some aspects of this common group of rusts. Figure 1 shows the upper surface of a hawthorn leaf with several infections of G. clavipes. The spermagonia can be seen as black spots within each of the colonies. In figure 2 a leaf of another hawthorn has been turned over to show an aecium of G. globosum. Figures 3 and 4 show telia of G. clavipes occurring on a juniper branch. In species of Gymnosporangium the teliospores are produced in long chains that form gelatinous columns called telial horns, most easily observed during wet weather. The telial horns can be seen in G. juniperi-viginianae (Figure 5) and G. nelsonii (Figure 6) arising from large spherical galls that the fungus has induced the plant to form.