Home >> Where fungi grow >> Mutualists >> Mutualisms with algae
MUTUALISMS BETWEEN FUNGI AND ALGAE
The term "algae" (singular: alga) is used to denote a vast array of plant-like organisms. Modern genetic studies have shown conclusively that the organisms called algae belong to several different kingdoms and are mostly not plants at all. We should probably abandon the term altogether, but it has a long history and is in fact not altogether useless.

The organisms we call algae are a diverse group of often very distantly related species. All of them lack roots and real leaves, although some of the larger ones have root-like structures called holdfasts and flat leaf-like blades. All contain chlorophyll and all are photosynthetic, but in many groups the green colour is masked by a variety of "accessory pigments". The smallest algae are one-celled and only a few micrometers in diameter. Nanochlorum eukaryoticum may be as small as 1 μm in diameter. At the other end of the scale, Nereocystis lutkeana, a giant "kelp" common on our west coast may be as long as 30 meters. In the photograph at right you can see the long leafy blades at one end of the thallus, which then snakes off to the right out of the picture.
LICHENS
Lichens are a mutualism formed between fungi and various groups of algae. The importance of this mutualism in terms of ecology, biodiversity and global well-being cannot be overstated. The most characteristic feature of lichens is that the combination of the two mutualists, alga (called the photobiont or phytobiont) and fungus (called the mycobiont), result in a form called a thallus (plural: thalli)that is completely unlike either of the two participants. The lichen thallus can be quite large and conspicuous, more closely resembling a moss or small plant than either a fungus or an alga.
The Thallus
Lichen thalli are generally classified into one of three types: crustose, foliose or fruticose. A fourth thallus type called squamulose is often recognized by lichenologists as well. Recognizing these basic types is not difficult.
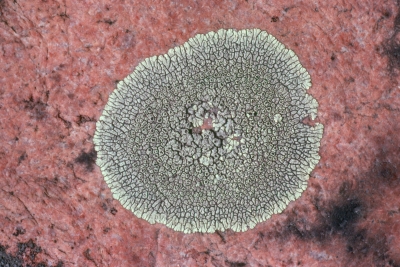
Crustose thalli are closely attached to their substrate so that they have a well-developed upper surface but simply merge into the substrate on the lower side. You cannot usually peel a crustose lichen away from its substrate nor will you see its margins curving back to reveal a compact lower surface. In the illustration at left the crustose lichen Dimelaena oreina is growing on the surface of a granite rock to which it is firmly attached. The places where it appears to have lifted off were probably caused by drastic forces, such as winter ice or being eaten by an animal.

Foliose lichens are like leaves (hence their name) and have a distinct upper and lower surface. In the picture at left a species of the foliose lichen genus Peltigera is growing on a mossy place on the forest floor. The ascending lobes make it look like a plant with leaves. In a few places you can see the nearly pure white underside of the lobes

Fruticose lichens are like small trees or shrubs and do nor exhibit an upper and lower surface; instead their branches have radial symmetry like a plant. If you cut one of their branches you will see that they may have an inner and outer surface, but never anything you would label top and bottom. The photograph at left includes two species, Cladina alpestris, the dense bushy one, and Cladonia rangiferina, the less densely branched one. Cladonia rangiferina, commonly called reindeer lichen, is a major source of food for caribou and reindeer during the winter.

Squamulose lichens are very similar to foliose lichens in having a two-sided thallus with a distinct top and bottom. Their main distinction is their small size; rarely being larger than 4 mm. They usually form closely-packed thalli together forming a shingle-like colony. The species of Cladonia at left is typical of its genus in producing not only a squamulose thallus but also upright structures called podetia that eventually support apothecia (see below for a further discussion of these structures). The cup-shaped structure in the photograph is a developing podetium.
Sexual reproduction
Most lichens are members of the Ascomycota and reproduce by means of asci and ascospores. For more information about these structures in fungi see the section on Reproduction in the Dikarya.

In the image at right the lichen Lecanora grantii, a common supralittoral species in our area, can be seen bearing several reddish brown, disc-shaped apothecia. Apothecia are structures that produce a palisade of asci that are openly exposed to the environment. Lichens producing apothecia are discussed further in the section dealing with lichen-forming discomycetes. In the case of L. grantii, the layer of asci and intervening tissues is reddish brown and is enclosed by the whitish margin of the apothecium. Although appearing whitish, the margin contains green algae, the photobiont, and will be seen to be green if cut lengthwise.

Some lichens produce their asci in perithecia such as those in the photograph of Hydropunctaria maura at right. Perithecia are flask-shaped structures that open to the outside by a pore called an ostiole. Lichens producing apothecia are discussed further in the section dealing with lichen-forming pyrenomycetes. The ostioles in both perithecia at right can be seen as small circular areas at their tops.


There are some other ways that asci can be produced by lichens, but perithecia and apothecia are by far the most common. The photographs above illustrate two other types you may encounter. The first are structures called lyrellae. These are similar to apothecia but are greatly elongated. In the case of Graphis scripta, shown in the photo, the lyrellae are highly branched and may resemble some kind of mysterious writing. The second photograph, of Calicium trabinellum, illustrates a mazaedium, a kind of stalked apothecium in which the asci dissolve and leave the ascospores to pile up in a powdery mass.
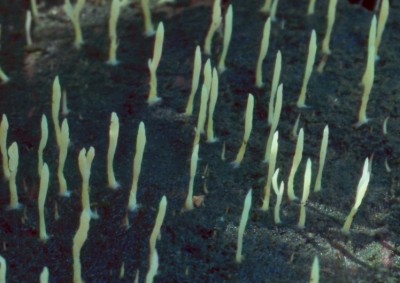

A few basidiomycetes are also capable of forming lichens. These are not generally considered to be highly-developed relationships yet there is no doubt they function as lichens. The first of the two photos above shows Multiclavula mucida. In this species the basidia and basidiospores line the surface of the upright "fingers" and under cool moist conditions release the spores to drift in the wind. The photobiont, a green alga, forms a thick crust of the the substrate, in this case rotten wood. The algae are enclosed by the hyphae of the mycobiont. In the second picture the mycobiont is Lichenomphalia umbellifera, a mushroom. The photobiont and its relationship with the phytobiont are the same as in M. mucida
Asexual reproduction
So far we have only discussed structures involved in the sexual reproduction of lichens. Since these sexual structures reproduce only the fungus, the resulting spores must be fortunate enough to land on an appropriate alga, or perish. However, there is another way. If the lichen can disperse propagules containing both myco- and photobionts then it will be able to develop in any suitable habitat. However, this type of reproduction is strictly clonal and does not allow for the kind of genetic recombination that occurs during sexual reproduction.


Clonal reproduction of lichens can occur in several ways. The simplest of these is simply to separate a piece of the thallus containing both alga and fungus and send it off by wind or water to develop in a new place. This kind of reproduction is common among lichens and generally effective. There are more highly developed forms of clonal reproduction, two of which are represented in the photographs above. In the first the lichen has produced soredia. Soredia are small bundles of algae held together by fungal hyphae. They are small enough to be carried by wind yet guarantee the presence of both partners. The illustration above left shows a young thallus of the foliose lichen Peltigera didactyla. In this species the upper surface becomes dotted with soralia, special structures for the production of soredia. In the photograph, the soralia have released granular masses of soredia. The other photograph above is a highly magnified view of isidia, small coral-like branches containing both mutualists that can break off and drift to a new habitat. The lichen in the picture is Xanthoparmelia conspersa, a common lichen on exposed rock in New Brunswick.
Lichen habitats
One of the fascinating aspects of lichen biology is the ability of these organisms to occupy habitats that would be totally in inhospitable to other organisms. Thus we can find them growing on the ground in deserts, on the sides of dry rock, hanging from the branches of trees and and even growing on the backs of turtles. They are nearly as easy to find and study in the middle of winter as during the warmer months.



The first of the three photographs above was taken in Saskatchewan, out in an open prairie. The rock in the forground is the highest point in the immediate area; animals sitting there get a panoramic view of the grassland and all that is taking place there. It is a favourite place for birds, especially birds of prey waiting for a mouse or vole that might be moving through the grass. The orange lichen is a species of Xanthoria that thrives on nitrogen-rich bird droppings left on the rock.
Similar species of Xanthoria, as well as members of the related genus Caloplaca, can be found on our seacoast on rocks frequented by gulls and cormorants. The second of the two pictures above is of White Horse Island, a small island in the Bay of Fundy supporting large colonies of nesting birds. The white colour of the rock is due to a thick layer of bird droppings; the orange material is a species of Caloplaca.
The third picture was taken at The Old Burying Ground in Kittery Point, Maine. The gravestone at left marks the resting place of Roland Thaxter (1858-1932), Professor at Harvard University and brilliant mycologist, known in particular for his monumental studies on the Laboulbeniales. Beside Roland's grave is that of his brother Karl. Both gravestones have become colonized by lichens and are now difficult to read. Click on the photograph to get an enlarged version of Roland's gravestone

Another interesting thing about our coastal lichens is that some of them are highly tolerant of salt, a substance that is toxic to most fungi, including lichenized ones. The picture at right depicts some coastal rocks on the Bay of Fundy near Saint John. At the bottom of the picture are bunches of brown algae, mostly Fucus vesiculosus and Ascophyllum nodosum, commonly called rockweed. These rockweeds grow in areas along the shore where they will be immersed in seawater, at least at high tide. At the very top of the rock is a patch of orange, probably Xanthoria parietina. In between is a black zone consisting of the custose lichen Hydropunctaria maura. Hydropunctaria maura can grow where it is periodically immersed in seawater but is also able to grow in an area just above that where it receives only splash from waves. This "black zone" occupies an area that often goes for days or even weeks without immersion in seawater but will eventually get splashed. This is a tough place to live: too saline for terrestrial organisms and too far above the sea for marine ones. Just the place for a lichen!

The picture at right depicts yet another species of Verrucaria mucosa, a close relative of H. maura. Unlike H. maura, V. mucosa grows where it gets innundated by seawater daily. In fact, it releases its ascospores when it is above the water and thus depends upon being exposed to air. However, it does not grow in the upper areas of the tide like H. maura does and can only be observed at low tide. In the picture V. mucosa can be seen as circular black spots resembling tar. On parts of the rock that have dried it is harder to see but you may notice that it is slightly green, revealing the presence of the photobiont. The red spots are the alga Hildenbrandia polytypa, similar is size and growth habit to V. mucosa but not lichenized and quite unrelated to it.

The last picture again shows Verrucaria mucosa, this time growing under water at high tide. Note that even this lichen has its limits; most of the rocks in the picture have no lichens at all. This may be because the rocks are too small and may be moved by currents as the tide ebbs and flows or it may be that their surfaces are unsuitable for lichens. Another problem that lichens face is being eaten by animals. Many contain acids and other compounds that make them unpalatable to animals but V. mucosa is not entirely immune. Notice the large rock above the one with lichens on it. On its surface is a small snail called a periwinkle. Some periwinkles, notably the rough periwinkle, eat V. mucosa and can remove it from its habitat. This has not happened here yet but there are in fact several periwinkles present, as well as the white barnacles and a mussel. How many periwinkles are here? Not many at first glance, but you might be surprised. Click on the picture to get an enlarged view and see how many periwinkles you can count.
THE MUTUALISM BETWEEN ASCOPHYLLUM NODOSUM AND MYCOPHYCIAS ASCOPHYLLI:
One of the more intriguing mutualisms found in our region is the one between the brown alga Ascophyllum nodosum and the fungus Mycophycias ascophylli. Ascophyllum nodosum, commonly called rockweed, occurs in the intertidal zone where it is left exposed to the air when the tide goes out. Mycophycias ascophylli, a member of the lichen-forming order of fungi Verrucariales, grows within the body (thallus) of A. nodosum where it is thought to help the seaweed resist drying out when it is exposed. In return the fungus has access to carbohydrates and other nutrients within its protective environment. Dr. D.J. Garbary and colleagues at St. Francis Xavier University in Nova Scotia have studied this mutualism in detail and have shown that the fungus not only forms relationships with the rockweed but also seems to form a mutualism with Polysiphonia lanosa, a common epiphyte found attached to the A. nodosum.

The pictures above show some aspects of the Ascophyllum/Mycophycias relationship. At far left is a thallus of A. nodosum spread out on a rock. You may wish to look further back on this page to see the habitat photo of A. nodosum. The next picture shows a detail from the first panel. The small almond-shaped structures along the stem are receptacles. Each receptacle bears a number of conceptacles, structures that release sperm and egg into the ocean each spring. These are seen as bumpy areas in the second photo but in the third more highly magnified panel they can be seen more easily and reveal the pores through which the sperm and egg escape. The next panel is even further magnified and the conceptacles are even clearer. In this panel it is also possible to see tiny black dots, resembling grains of pepper; these are the perithecia (fruiting bodies) of Mycophycias ascophylli. The blue box drawn on around one of these leads to the next photograph, taken with a compound microscope, showing a detailed view of one perithecium partially submerged in the receptacle. The perithecia contain asci and ascospores. The last panel shows one ascus containing eight 2-celled ascospores. The ascospores are not very clear in this picture but are nevertheless nearly mature.

In our region Ascophyllum nodosum releases its sperm and eggs in late May. On a warm day at low tide these tiny cells ooze out of the conceptacles like toothpaste out of a tube. When the tide comes in they are released into the water. The sperm, released in numbers large enough to colour the water orange, swim activly in search of eggs and attach to them when they find one. Eggs can be observed spinning wildly, powered by hundreds of sperm attached to their surface. Finally one sperm succeeds in fertilizing the egg, which then sinks to the bottom to grow into a new plant. At the same time sperm and eggs are being released the asci of Mycophycias ascophylli shoot their spores into the air. It is likely that these spores are able to colonize the newly developing plants but the details of this stage of development are still not very well understood. The orange colour in the water caused by massive numbers of sperm and eggs could be mistaked for a toxic "red tide", but that is a completely unrelated phenomenon.
There has been some debate about the Ascophyllum/Mycophycias relationship. Most biologists accept Dr. Garbary's contention that it is mutualistic but don't always agree about its significance. Some have suggested that this is a type of lichen, arguing that a mutualism between an alga and a fungus is by definition a lichen. However, others have argued that in a true lichen the fungus is the exobiont, that is it encloses the alga and forms the conspicuous part of the thallus. In the Ascophyllum/Mycophycias mutualism the fungus is clearly the endobiont and contributes little to the appearance of the alga. Recent genetic evidence that Mycophycias ascophylli is related to the lichen-forming Verrucariales lends support to the lichen proponents.